Malá škola kaktusáře
MUDr. Vladimír Plesník
Areoly a postavení trnů
Původ areol a trnů byl u kaktusů dlouho nejasný. Teprve později se zjistilo, že trny jsou vlastně přetvořené větvičky s listy a že představují růstové vrcholy. Dolní část areoly bývá zpravidla zvětšena, takže růstový bod (vegetační vrchol) bývá uložen při jejím horním okraji. Část areoly, z níž vyrůstají trny se vyvíjí zdola nahoru, proto také trny vyrůstají postupně od dolní části areoly směrem nahoru. Na protáhlých areolách dochází k hřebínkovitému (pektinátnímu uspořádání trnů. U některých rodů (např. Coryphantha, některých cereusů) vzniká mezi vegetačním vrcholem a dolní částí areoly, nesoucí trny, rýha tzv. sulcus.
Diagram areoly:
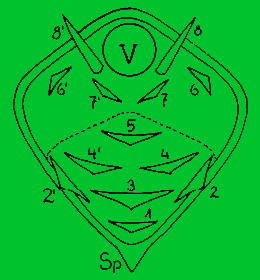
V - růstový vrchol areoly
Nárys květuschopné areoly Leuchtenbergia principis.
Čísla označují postupný růst trnů. Semenáčky mají trny č.1 - 3, na starších, ale nekvetoucích areolách, jsou trny č. 1 - 5
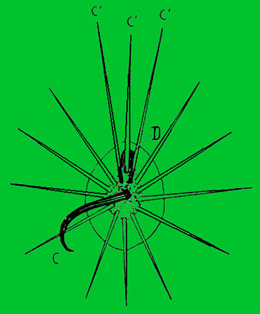
D - nektarové vyústění žlázy
Areoly Hamatocactus setispinus se středními (C a C´) a paprsčitě uspořádanými trny.

Areola z temene Encephalocarpus strobiliformis, mající ještě 3 nektarové trny
Areola Cumarinia odorata se 3 háčkovitými trny a rýhou
Areola Mammillaria candida, vnitřní trny jehlovité, ostatní štětinovité
Areola jednoho druhu Neolloydia, s hřebenovitě uspořádanými trny
Areola Tephrocactus papyracanthus s papírovitě rozšířenými trny a s glochidou
Areola Ferocactus latispinus s šídlovitými trny a jedním hákovitým trnem
Trny vyrůstají vždy od spodu (base), proto jsou zde nejměkčí. Jejich špička vysycháním tvrdne. Vyjímkou jsou některé rody (Hamatocactus, Ferocactus, Coryphantha, některé Opuntia), jejichž některé trny se stále pomalu prodlužují. Zůstávají pak poněkud šťavnaté a na jejich povrchu se vylučuje sladká, lepkavá šťáva (nektar – proto název nektarové trny). Zvlášť nápadné jsou u epitelant a encefalokarpusu. U těchto rodů vyrůstají z horní části mladých areol zvláště dlouhé trny, vylučující na špičce vodnatou tekutinu. Mezi trny, chlupy a vlasy nejsou podstatné rozdíly. Vlna v areolách je však produktem buněk pokožky.
Glochidy se skládají většinou z jemných, hustě uspořádaných trnů, jejichž báze jsou trvale měkké a lámavé, kdežto povrch jejich ztvrdlých špiček tvoří šupinatě odstávající buňky. Proto trny z glochid po zapíchnutí do kůže tak dobře v ní drží. Podobnou úpravu povrchu mají i silnější trny některých opuncií. Délka, síla a vybarvení trnů se mění v rámci jediného druhu, od rostliny k rostlině, někdy i na téže rostlině, dojde-li ke změně pěstebních podmínek. Stejně se mění i počet trnů v areole. Jejich uspořádání – postavení v areole, tzv. diagram areoly, je stálý a často umožňuje soudit na příbuzenskou souvislost rostlin. Postupem doby si kaktusáři vytvořili vlastní názvosloví pro odlišně vyhlížející trny, poněkud však chybí jednotný, internacionální výklad užívaných pojmů. V příští kapitole se pokusím přinést přehled základních názvů trnů spolu s jejich nákresem, který bývá instruktivnější než vlastní popis.
Vzhled a názvy trnů

Šídlovitý: u base široký, vybíhající do tenké špičky (subulate)
Kuželovitý: u base široký, krátký (conic)
Jehlovitý: u base tenký, vybíhající do tenké a dlouhé špičky (acicular)

Zprohýbaný: nepravidelně pokroucený (tortuous)
Kuželovitý: špička je stočena zpět (uncinate)
Jehlovitý: tenký, tuhý, vlasovitý (bristles)

Pochvovitý: povlečený tenkou papírovou vrstvou (sheathed)
Glochidy: krátké štětiny s háčky (glochids)
Prstencovitý: na povrchu má vroubky (annulate)

Kolmo odstupující: (porrecte)
Pektinátní: hřebínkovitě uspořádané
Zpětně zahnuté: špička směřuje dolů a k tělu rostliny (appressed)

Odstupující: rovné trny směřující špičkou šikmo dolů (deflexed)
Střední vystupující téměř ze středu areoly, krajní z okrajů
Přilehlý: přiléhá k žebru nebo mezižeberních rýh (appressed)
Typy a vlastnosti kořenů
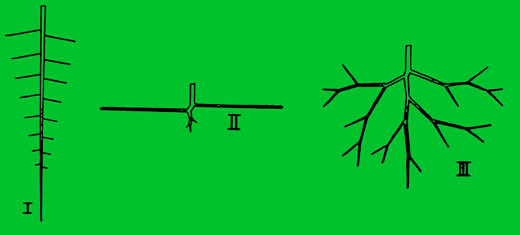
I. Kůlový kořen, II. Ploše rozvětvený kořen, III. Srdčitý kořen

Řepovitý kořen - Hlízovité kořeny
O charakteristickém vzhledu a vlastnostech kořenů se toho ví poměrně málo. Důvodem je to, že jejich studium lze uskutečnit prakticky jen na nalezištích.
U nás se kaktusy pěstují zčásti roubované, nebo se pikýrují či pěstují v květináčích, což ovlivňuje jejich přirozený růst kořenů. Kaktusy tvoří hlavní kořen, směřující z osy rostliny do středu země a postranní kořeny. Mimo nich jsou dále adventivní (přídatné) kořeny, vedoucí později k větvení kořenů. Na odnožích a zvláště u epifytních kaktusů vznikají „odnožové“ kořeny, mezi něž patří také „vzdušné“ kořeny. Tyto odnožové kořeny se tvoří v hloubce těla odnože z cévního svazku a musí nejdříve proniknout navenek. Při zakořeňování rostliny, která z nějakého důvodu ztratila původní kořeny, brání velmi často hrubá korkovitá tkáň na kořenovém pahýlu proniknutí nových kořenů. Doporučuje se proto změkčit tuto vrstvu teplou vodou, tupým nástrojem ji rozrušit a opatrně odstranit. Na 2 – 3 týdny ponecháme rostlinu v suchu, aby se rány bezpečně zahojily, pak ji znovu vysadíme. Zejména u importů bez dobře vyvinutých kořenů je vhodné odříznout zbytky kořenů až na zdravý cévní svazek a rostlinu zakořeňovat jako odnož.
Troll rozlišuje tři základní typy kořenů:
- Kůlový – dlouhý, s relativně krátkými postranními kořeny, u některých rodů je přetvořen na řepovitý kořen.
- Ploše větvený – krátký, se silnými a dlouhými postranními kořeny, které odstupují v malé hloubce
- Srdčitý – tvoří přechod mezi předchozími typy, na průřezu jsou kořeny rozloženy do srdčitého tvaru
Hlízovité kořeny se liší od řepovitých tím, že u řepovitých má „řepa“ funkci hlavního a tvar kořene, kdežto hlízy vznikají na postranních kořenech a mají funkci zásobních orgánů. Nejznámější kořenové hlízy má např. Ancistrocactus megarhizus, řada wilkoxií, opuncií a další.
Nejčastěji nalézáme u kaktusů kořeny ploše rozvětvené, mnohdy do značných vzdáleností. Britton a Rose sledovali délku postranních kořenů u Carnegiae (17 – 20 metrů), u Melocactus intortus v hloubce pouhých 5 cm dosahovaly kořeny délky 7 metrů. Na nalezištích bývá ve svrchních vrstvách půdy největší koncentrace živin. Široce rozvětvený kořenový systém umožňuje kaktusům mimořádně bohatou nabídku živin. Vysazování kaktusů do květináčů podstatně omezuje růst jejich kořenů. Proto mnohem lépe rostou voně vysazené rostliny, skupinovitě vysazené do velkých misek.
Při správném pěstování tvoří se kořeny na bazální (spodní) části kaktusu, z nejspodnějších internodií (vysvětlím později). Kořeny vznikající na odnožích dále nerostou, pokud odnož zůstává na mateřské rostlině, která je živí. Pravé vzdušné kořeny vznikají z vyšších těla především u epifytních a šplhavých druhů kaktusů. Jejich úkolem je jak výživa rostliny, tak přichycení k podkladu. S tvorbou vzdušných kořenů se setkáváme i u cereusů (např. Trichocereus macrogonus, schickendantzii). Příčinou bývá někdy poškození kořenů (hniloba, kořenovka), příliš malý květináč, nedostatek vláhy nebo živin, ale také zimování těchto rostlin bez půdy a vleže.
U notokaktusů nacházíme na kořenech odnože, pocházející z dolních areol těla a rostoucí zpočátku pod svrchní vrstvou půdy. Protože tyto odnože nemají chlorofyl, bývají začátečníky omylem pokládány za kořenové „nádory“.
Kořeny, které si rostlina vytváří při hydroponickém pěstování, jsou odlišné stavby než kořeny vznikající v půdě. Proto náhlý přechod z hydroponie na pěstování v substrátu vede téměř vždy k zástavě růstu ne-li ke ztrátě rostliny.
K urychlení růstu kořenů při zpravokořeňování roubů a odnoží se dnes u nás s úspěchem používá stimulační pudr VS 9, kterým se řezná rána lehce popráší a nechá zasychat. Pokud je pudr čerstvý a správně skladovaný, spolehlivě vede k tvorbě silného svazku kořenů do 10 dnů.
Tvary a názvy těl kaktusů - (část 1.)
Tělo kaktusu je tvořeno zeleným zdužnatělým stonkem, jehož kulovitý nebo válcovitý tvar poskytuje rostlině co největším objem tekutin při co nejmenší možnosti její ztráty výparem z povrchu rostliny. Uvnitř je osové pletivo poskytující stonku pevnou oporu a současně růstovou osu. Při její poruše vznikají nepravidelná – monstrosní těla, nebo hřebenité útvary – kristáty.
Cereusy jsou kaktusy s tělem sloupovitým (kolumnárním), válcovitým (cylindrickým), nebo šlahounovitým. Jednotlivé šlahouny mohou být vzpřímené (trsovitý – cespiotosní růst, obr. 1), nebo poléhavé (některé haageocereusy) či převislé (např. aporokaktusy, rhipsalisy). Někdy se hlavní stonek cereusu v určité výšce osy větví a vznikají tvary stromovité (obr. 2) nebo kandelábrovité (obr. 3).

- Trsovitý růst cereusů
- Stromovitý růst cereusů (arborescentní)
- Kandelábrový růst
Název kaktoidní kaktus označuje rostliny s kulovitým tělem, které může být protaženo až v krátce válcovité tvary, nebo naopak stlačeno na ploché až terčovité tvary (např. Astrophytum neboGymnocalycium asterias).
Odnožující rostliny vytváří shluky hlaviček. Také takovému růstu říkáme trsovitý. U některých rostlin (např. Mammillaria parkinsonii) se hlava dělí na dvě souměrné části, která se časem opět stejným způsobem dělí dále (dichotomické dělení – obr. 4 a – c).

Postup dichotomického dělení hlavy mamilárie
Tvary a názvy těl kaktusů - (část 2.)
Řada kaktusářů začínala s pěstováním opuncií, někteří jsou jim věrni stále. U nás asi není specializovaná sbírka opuncií, přesto řada z nich si zaslouží naši pozornosti. Články opuncií mají tyto základní tvary:

Okrouhlý (orbicular): plochý článek má téměř kruhový tvar, připomínající list nebo květní plátek
Opakvejčitý (Obovate): jako vajíčko postavené na špičku. Nový článek se ke staršímu připojuje vždy užším koncem

Zoubkovaný (repand): okraje článku hrubě zoubkované, často i zprohýbané
Protáhle oválný (oblong): délka článku je 2 – 3x větší než jeho šířka

Oválný (repand): (elliptical): typické je, že „zašpičatění“ na obou koncích článku je stejné
Protáhlý (elongated): v poměru k šířce je článek značně delší až válcovitý
Příklady rostlin: okrouhlý = Opuntia setispina, opakvejčitý = Opuntia affinis, protáhle oválný = Opuntia maxima, zoubkovaný = Opuntia dillenii, oválný = Opuntia ficus-indica, protáhlý = Opuntia elatavar. elongata.
Stavba semen
Slupka semene (osemení, testa) má dvě vrstvy. Zevní může být hladká a tvrdá nebo měkká, bradavčitá nebo dolíčkovaná (viz. Jub. sborník str. 18 – 34).

Semeno Echinocactus saltilensis, pohled za strany hilu a pohled z boku

Hilum je místo, kde se na semínko připojuje semenný provazec „pupečník“. Často je poblíž hilu také více či méně zřetelný otvůrek – mikrophyla.
aJ – zevní testa, P – perisperm, iJ – vnitřní testa, E – embryo, Esp – endosperm, Mi – mikrophyla, Hi – hilum „pupek“.
 Semeno Thelocactus bicolor - kráterovité hilum
Semeno Thelocactus bicolor - kráterovité hilum
 Echinomastus unguispinus
Echinomastus unguispinus
 Lobivia wrightiana - hilum. F – jizva po provazci (funikulum)
Lobivia wrightiana - hilum. F – jizva po provazci (funikulum)
 Mammillaria longiflora
Mammillaria longiflora
 Astrophytum myriostigma – čepicovité hilum
Astrophytum myriostigma – čepicovité hilum
Vnitřek hilu bývá často uzavřen jen tenkou korkovitou vrstvou, která se při klíčení trhá. Někdy je hilus vyplněn silnější korkovitou vrstvou, označovanou jako strophiola nebo arillus. Tato vrstva může i přesahovat okraj osemeníku. Typická je strophiola pro mamilárie z podrodu Phellosperma.
Po opylení dojde ke spojení jádra pylu (spermatu) s vaječnou buňkou. Oplodněné vajíčko se mění na zárodek (embryo). V oplodněném vaječném lůžku se buňky také množí a vytvoří endosperm, sloužící k výživě embrya. U kaktusů bývá endosperm spotřebován jakmile se počne rozvíjet embryo. Při dozrávání semene se změní zevní blanka (integument) na pevnou zevní vrstvu testy. V místě připojení semenného provazce (funiculum) vzniká pupek – hilum. Zásobu živin pro zárodek obsahuje jak endosperm, tak perisperm.
Stavba a vývoj semenáčku

Stav po vyklíčení semene

Vznik pupene epikotylu mezi děložními lístky
Se stoupající sukulencí rostlin se zmenšují děložní lístky vyklíčených rostlinek a zvětšuje se hypokotyl. U vysoce sukulentních rostlin jsou děložní lístky redukovány až na malé hrboly vyrůstající v horní části hypokytylu.
Prvé základy listů – podaria, jsou na semenáčcích uspořádány zpravidla spirálovitě. Na malých semenáčcích není však uspořádání těchto základů listů zvláště výrazné. Proto na nich vidíme volně a jednotlivě rozložená podaria v podobě bradavek

Další vývoj sukulentního epikotylu mezi děložními lístky

Epikotyl (areoly vyrůstající v ose v děložních lístků)
U druhů s malým počtem základů listů velmi brzo začínají podaria splývat a tvoří tak žebra. Mladé rostlinky mají obvykle menší počet žeber. Tento se zvyšuje až při dalším růstu rostliny.
Nedostatečná výživa nebo osvětlení zvláště při česném vývoji rostlin (ale i později) může vést ke snížení počtu žeber.
Vzácně dochází k určitému posunu v postavení podarií. Vzniká tak nápadný šroubovitý růst žeber. Při úplném rozrušení symetrického uspořádání podarií vzniká nepravidelný – monstrózní růst.
Některé botanické názvy
Značná část kaktusářské literatury, která je u nás dostupná, je psána německy. Pokládám proto za vhodné, současně uvádět i německé názvy.
A. Útvary na pokožce rostlin:
- trny (Dornen) = přeměnné listy nebo větvičky
- ostny (Stacheln) = tvrdé, pichlavé výrůstky na pokožce
- vlasy (Haare) = jedno nebo více buněčné, jednoduché nebo rozvětvené, rovné nebo stočené výrůstky z pokožky rostliny
- štětiny (Bortshaare) = tuhé a pichlavé
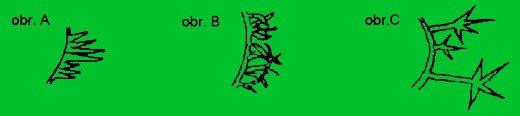
- nektarové vlasy – brvy (Drüsenhaare) = vlasy mající na špičce nektarové žlázy (viz obr. A)
- plsťové vlasy – brvy (Kraushaare) = tuhé, delší zkroucené (obr. B)
- hedvábné vlasy – brvy (Seidenhaare) = husté, přiléhavé, lesklé
- hvězdicovité (Sternhaare) = hvězdicovitě se rozvětvující (obr. C)
- vlna (Wollhaare) = měkké, husté, dlouhé vlasy
- ojíní (Rief) = bělavý nebo namodralý povlak pokožky, který lze setřít
B. Tvary okvětních plátků:

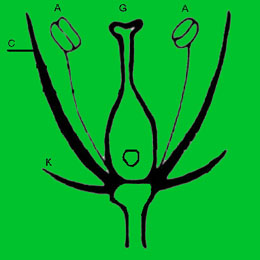
C. Části květu:
- květní obaly (Blütenhülle) = zevní nepohlavní části květu, sestávající z volně stojících nebo srostlých okvětních plátků, tvořící tak trubku. Zevní okvětní plátky se nazývají kalich, vnitřní pak – koruna.
- kalich (kelch) = se skládá z většinou zelených, volných i srostlých kališních lístků
- koruna (Krone) = se skládá z barevných, volně nebo částečně srostlých okvětních plátků
- okvětní plátek (Kronblatt) = je část koruny, obvykle mající užší část, tzv. nehet (Nagel) a širší část (Platte)
- prašníky se skládají z tyčinek – prašníkových nitek (Staubfaden) a prašníkových váčků (Staubbeutel) obsahující pyl.
- ovarium (Fruchtebeutel) obsahuje zárodky semen, několik ovarií srůstá a tvoří semeník (Fruchtknoten), z něhož vyrůstá čnělka (griffel) nesoucí různě utvářenou bliznu (Narbe).
- jednopohlavní květy = mají buď prašníky (mužský typ) nebo ovária
- oboupohlavní květy = mají prašníky i ovaria
- jednodomé rostliny = na téže rostlině rostou mužské i ženské typy květů
- dvoudomé rostliny = na rostlině rostou jen „mužské“ nebo „ženské“ typy květů.
Skladba květu
Květ, latinsky Flos, řecky Anthos, je rozmnožovací orgán rostlin vytvářejících semena. Vzniká přeměnou listů a skládá se ze samčích tyčinek a samičích pestíků. Tyto jsou obklopeny květními obaly v podobě kalicha a korunky nebo okvětí.
Květ může být obojaký – má tyčinky i pestíky, nebo jednopohlavní – jsou v něm jenom tyčinky nebo jen pestíky (Euphorbia obesa, Mam. dioica aj.)
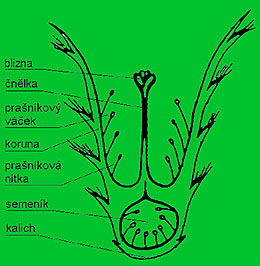
Soubor tyčinek (androceum) se skládá z jedné až několika tyčinek (stamen), které jsou samčím květním ústrojím, vytvářejícím pyl.
Tyčinka se skládá z nitky a prašníku (anthera), který má dva pylové váčky.
Pestík (pistillum) se skládá z blizny (stigma), čnělky (stylus) a semeníku (ovarium). Blizna bývá lepkavá, chlupatá, někdy členěná na několik laloků. Někdy je pestík delší než tyčinky, u jiných rostlin je tomu naopak. Tím má být zabráněno samoopylení.
Kalich (calix) je zelený vnější obal květu, tvořený volně rostoucími nebo srostlými lístky. Může mít různé tvary.
Koruna (corolla) je vnitřní, zpravidla zbarvený obal. Korunní plátky bývají volné nebo srostlé, různého tvaru.
Ke zkrácení a zjednodušení popisů květů se často užívají květní diagramy. V psaném diagramu se užívají tyto stálé značky:
| P | okvětí (perigonium) |
| K | kalich (calix) |
| C | koruna (corolla) |
| A | tyčinky (androceum) |
| G | pestíky (gyneceum) |
 |
oboupohlavní květ |
 |
tyčinkový samčí květ |
 |
pestíkový samičí květ |
 |
souměrný květ |
 |
pravidelný květ |
 |
velký počet |
 |
svrchní semeník |
 |
spodní semeník |
| (3) | uvedený počet plátků nebo tyčinek příslušného kruhu spolu srostl |
Fyziologie rostlin
Fyziologie studuje látkové složení látkovou výměnu a přeměnu růst, dráždivost, pohyb rostlin a jejich rozmnožování a sleduje jejich závislost na životním prostředí.
Chemické složení rostlin, látková výměna
Zastoupení jednotlivých prvků v rostlinách se mění, některé jsou přítomny ve všech rostlinách ve značném množství, jiné jen ve stopách. Patří mezi ně uhlík, kyslík, dusík, vodík, síra, fosfor, draslík, vápník, železo, a hořčík. Ty prvky, které bývají přítomny ve větších množstvích, označujeme jako makroelementy (mimo výše uvedených sem někteří řadí i sodík, chlor, hliník a lithium). Dále jsou pro některé rostliny nezbytné prvky přítomné aspoň v malých množstvích, tzv. mikroelementy (zinek, brom, mangan, měď, jod, arsen, bor, fluor, olovo, titan aj.). V nadbytku však mohou tyto prvky působit škodlivě.
O významu jednotlivých prvků pro růst a život rostliny se můžeme přesvědčit pokusně. Užívá se k tomu pískových kultur, které se zalévají roztokem živin o přesně známém složení. Ještě jednodušší je vodní kultura, známá z hydroponie.
Význam prvků pro rostliny
- Dusík je nutný k tvorbě rostlinných bílkovin. Ve vodní kultuře nedokážou rostliny využít dusík ze vzduchu (chybí bakterie přítomné v půdě). Dusík rostlina přijímá v podobě roztoků dusičnanů, dusinatů a amonných solí, jeho nedostatek vede k zakrnění těla.
- Síra je stavební součástí bílkovin rostlinného těla. Při jejím nedostatku rostliny zpomalují růst a žloutnou. U kaktusů je známo, že některé druhy nutně vyžadují síru v podobě sádry (utahie, sklerokaktusy, některé koryfanty a mamilárie aj.).
- Fosfor je důležitou složkou bílkovin obsažených v jádře rostlinné buňky. Při nedostatku jsou rostliny bledé, nekvetou, málo odolné.
- Draslík – jeden z nejvýznamnějších prvků, potřebný pro látkovou výměnu k tvorbě cukrů a bílkovin. Nedostatek vede u listnatých stromů ke vzniku malých, rychle opadávajících listů.
- Vápník zvyšuje odolnost rostlin proti suchu a chladu, listy jsou při nedostatku vápníku bledé. Má snad vliv na kvalitu ostnů.
- Hořčík je nutný pro tvorbu rostlinného barviva – chlorofylu.
- Železo má stejný význam pro tvorbu chlorofylu a pro výměnu plynů v tkáních rostliny.
- Uhlík je nejdůležitější ze všech biogenních prvků. Bez uhlíku nemůže existovat žádná živá buňka. Rostliny jej přijímají ze vzduchu v podobě kysličníku uhličitého. Proto vyšší obsah kysličníku uhličitého ve skleníku bývá často nepoznanou příčinou lepšího růstu rostlin v tomto prostředí.
- Vodík – účastní se na stavbě všech molekul živé hmoty. Rostliny jej získávají z vody, svůj význam má i nepatrné množství – stopy tzv. těžkého vodíku, který podněcuje růst rostlin.
- Kyslík přijímají rostliny jen ze vzduchu v podobě kysličníku uhličitého. Nadbytek kyslíku vyvolává u rostlin „narkózu“ – zpomalení životních pochodů. Existuje však velká variabilita mezi jednotlivými rody.
Buňka a její činnost
Rostlinná buňka je buď částí vyšších rostlin, složených z mnoha buněk ale může sama o sobě představovat celý organismus (např. u mikrobů). Pro život buňky je nezbytná voda. Ta také tvoří největší část váhy rostlin i člověka. Množství vody v rostlině závisí na jejím druhu, stáří a nabídce vody. Vodní rostliny obsahují až 98% vody, okurky a salát 98 – 96% vody, brambory 75% vody, dřevo našich stromů asi 45% vody. V semenech bývá vždy méně vody a teprve dodání vody vede ke klíčení.
V buňce ani jinde v těle rostliny nebývá obsažena čistá voda Vždy je v ní obsaženo určité množství rozpuštěných látek, někde více, někde méně. Jsou – li tyto rozdílně koncentrované roztoky od sebe odděleny polopropustnou blanou, mohou jí procházet jen molekuly vody, ne však rozpuštěná látka. Vždy voda proudí z roztoku řidšího do koncentrovanějšího a to tak dlouho, až dojde k vyrovnání koncentrací. Tomuto jevu se říká osmosa.
Buněčné blány a tenká vrstvička na špičkách kořínků jsou také polopropustné. Voda proniká z půdy nebo okolí do buněk jen tehdy,je – li v buňce koncentrovanější roztok. Čím je půda sušší, tím větší sací sílu musí rostlina vyvinout, musí mít tedy vyšší osmotický tlak. Bahenní rostliny mívají nízký osmotický tlak, kdežto suchomilné rostliny včetně kaktusů mají obrovský osmotický tlak. Při nadbytku vody se buňky suchomilných rostlin trhají, tělo rostliny praská a tato hyne.
Osmotický tlak uvnitř buňky, který pružně rozepíná buněčnou stěnu se jmenuje turgor. Průduchy mezi buňkami se otevírají jen tehdy, je – li turgor velký. Při ztrátě většího množství vody rostlina vadne a průduchy se zavírají. U kaktusů je otevírání průduchů a hospodaření s vodou ještě komplikovanější.K této problematice se ještě později vrátíme.
Roztok s vyšším osmotickým tlakem je hypertonický, s nižším je hypotonický, jsou li oba roztoky se stejným tlakem, říkáme že jsou isotonické. Zálivka koncentrovanějším roztokem živin je hypertonická a nemůže být rostlinou využita.
Proudění vody v rostlině ovlivňuje především osmosa, vztlak kořenů, kohese a kapilarita vody a její odpařování – transpirace.
Kořenový vztlak můžeme pozorovat při roubování na dobře napitou podložku:z cévního kroužku vystupují kapičky šťáv. Příčinou vztlaku je rovněž osmosa. Kohese je název pro síly, které mezi sebou vážou molekuly jakékoliv látky. Kohese molekul kamene, železa či dřeva je vysoká, kohese vody je malá, ale patrná. Voda proudící v tenoučkých cévách „kapilárách“ se v pokožce nebo v listech odpařuje. Sloupeček tekutiny v kapiláře se neustále posunuje k místu odpařování. Odpařování čili transpirace slouží rostlině jak k přenosu živin z kořání do celého těla, tak k ochlazování. Odpařením 1 g vody se rostlina zbaví 539 cal. tepla. U kaktusů je transpirace vody omezena na minimum a tento způsob ochlazování je poměrně málo využíván.
Mnohé látky potřebné pro rostliny se však ve vodě nerozpouští. Proto rostliny svými kořínky vylučují do okolí kyseliny, kterými rozleptávají nerosty. Pro čerpání živin z půdy je velmi důležitá reakce půdy. Většina kaktusů dobře roste v půdě neutrální nebo slabě kyselé (pH 6 7,5). Epyfitní kaktusy, např. Zygocactus, Epiphyllum a další, lépe rostou v kyselejší půdě o pH 5,5. Naopak některé vápnomilné rody z Mexika (Utahia, Sclerocactus aj.) vyžadují zásaditou reakci půdy (pH 7,5 – 8).
Látková výměna a dýchání
Látková výměna (metabolismus) rostlin se skládá ze dvou pochodů: asimilace a disimilace.
Asimilace je v podstatě příjem cizích látek (živin) a jejich přeměna na rostlinné tkáně. Pro všechny rostliny, stejně jako pro všechen život na světě, je nejvýznamnějším způsobem asimilace tzv.fotosyntéza. Při ní zachycuje zelené barvivo rostliny „chlorofyl“ sluneční energii. Pomoci chlorofylu a sluneční energie pak rostlina mění vodu a kysličník uhličitý, získávaný z okolního vzduchu, na cukry. Přebytečný kyslík vznikající při fotosyntéze je rostlinami uvolňován do vzduchu.
Mimo chlorofylu spolupůsobí při fotosyntéze i další rostlinná barviva – oranžové karotenoidy a žlutý xanthofyl. Tato zachycují ze slunečního světla část paprsků modré a zelené barvy, kdežto chlorofyl zachycuje žluté a červené spektrum barev. Protože v řadě rostlin (i v kaktusech převládá chlorofyl, je osvit s větším podílem žluté a červené složky účinnější.
Výsledkem chemických reakcí vyvolaných sluneční energií je vznik nejprve jednoduchých cukrů, pak složitějších látek, aminokyselin, bílkovin a tuků. Světlo je nezbytné pro zahájení fotosyntézy, řada dalších reakcí probíhá i za tmy.
Asimilace se však týká i příjmu dusíku, draslíku, fosforu, vápníku a jiných nezbytných živin. Rostliny je přijímají hlavně v podobě sloučenin s dusíkem, které přetváří na aminokyseliny a bílkoviny. Mimo nich vzniká v těle rostlin řada více méně složitých sloučenin, vitamínů, alkaloidů, rostlinných barviv a dalších.
Rychlost a účinnost fotosyntézy ovlivňuje zejména přítomnost vody, živin, dostatek světla, vhodná teplota a koncentrace kysličníku uhličitého.
Disimilace (dýcháni) je opakem fotosyntézy. Rozkladem cukrů dochází k uvolňování energie. Ta pak slouží k úhradě energie na životně důležité růstové a rozmnožovací pochody.
Zatím co ve dne převládá asimilace, za tmy dochází k disimilaci. Podobně jako pro fotosyntézu je nezbytným předpokladem přítomnost slunečního světla, je i pro disimilaci zapotřebí zvláštních působků –katalyzátorů. Za jejich pomoci probíhá rozklad nastřádaných zásob. Tyto katalyzátory bývají označovány jako enzymy. Většina z nich má úzce specifický účinek. Uplatňuje se pouze při jediné reakci a jen za určitých podmínek. Počet těchto katalyzátorů v těle rostliny je neobyčejně velký. Odhaduje se, že v těle nejjednodušších rostlin – bakterií - je jich asi 1000. Nejintenzivněji probíhá disimilace v mladých, vyvíjejících se částech rostlin. Zejména jsou to klíčící semena, bouřlivě se vyvíjející semenáčky a některé části květů.
Výměna asimilátů mezi rostlinami je základem tzv. symbiózy. Ve vzájemně výhodném vztahu žijí mnohdy i značně vzdálené dvě rostliny. Např. motýlokvěté rostliny vylučují některé asimiláty kořeny. Zde jsou využívány části půdních bakterií, které naopak obohacují okolí kořenů o dusíkaté látky neboť dovedou využívat vzdušný dusík.
Jen zřídka se pomýšlí na symbiózu kaktusů s jinými rostlinami. V popisech nalezišť bývá jen povšechně uváděna průvodní flora, složení půdy bývá zkoumáno jen z hlediska fyzikálního a chemického. Není však vyloučeno, že mimo struktury půdy, jejího pH, provzdušnění atd. se zde uplatňují i dosud neznámé, symbioticky žijící drobnohledné rostliny.
Růst, rozmnožování, pohyby rostlin
Růst rostlin je spojen s tvorbou buněčné plasmy a s dělením buněk. Vyšší rostliny rostou pouze ve vegetačních vrcholech a v obdobích, které se stále střídají s obdobím klidu. S růstem úzce souvisí vývoj rostliny. Po dosažení určité velikosti rostlina kvete a přináší plody. Často se stává, že dospělé rostliny nechtějí kvést, ač mají zdánlivě nejpříznivější podmínky k životu. Bývá to obvykle proto, že předchůdci této rostliny žili v prostředí s odlišnými světelnými, teplotními či dalšími podmínkami.
Rozmnožování rostlin může být pohlavní a nepohlavní. Pohlavním rozmnožováním označujeme sprášení blizny květu jedné rostliny pylem jiné rostliny téhož druhu (zřídka odlišného druhu). Nasazení plodu a vznik semen představuje ukončení tohoto pohlavního cyklu.
Nepohlavní, tzv. vegetativní rozmnožování, má mnoho forem. Patří mezi ně tvorba výtrusů nebo pučení (u výtrusných rostlin), plazivé osy (např.jahody), oddenky kdy z každé odložené části vyrůstá nová rostlina (pýr, přeslička, konvalinka), hlízy a cibule (tulipány, sněženky aj.), adventivní kořenové pupeny (švestky) a řízky (množení pelargonií, fuchsií, ale i růží a stromů). U kaktusů představuje typickou formu vegetativního množení, vznik odnoží, vyrůstajících z areol. U některých notokaktusů se tvoří podzemní odnože („brambůrky“) u kořenového krčku. Vegetativně množené rostliny mají všechny své vlastnosti naprosto shodné s mateřskou rostlinou. Tím se liší od semenáčků, kde dochází ke kombinaci vlastností obou rodičů. Roubování samo o sobě není množením. Vzniklé odnože na zbytku rostliny po odřezání „hlavy“ však mohou později sloužit k vegetativnímu množení. U rostlin, které nemůžeme sprášit, nebo kde je výsev velmi obtížný, užívá se umělého poškození vegetačního vrcholu k tomu, aby jsme rostlinu donutili k odnožování a tak získali množitelský materiál.
Rostliny, stejně jako zvířata či člověk, reagují na vliv prostředí. Tomu se říká dráždivost. Odpovědi na vnější podnět bývá obvykle pohyb. Podráždění může vyvolat vzájemný dotyk rostlin, světlo, elektřina, gravitace, chemické látky nebo teplo.
Reaguje – li rostlina na podráždění pohybem celého těla, říkáme tomu taxe. Např. fototaxe je naklánění těla rostlin za světlem, dobře známé pěstitelům kaktusů za oknem. Reaguje – li pouze některá část rostliny, mluvíme o tropismu. Např. růst kořenů vždy ve směru zemské přitažlivosti je tzv. geotropismus. Tigotropismus je reakce na dotyk – ovíjivé úponky popínavých rostlin. Fototropismus je dokumentován orientací postavení listů ke světlu nebo zešikmení temínka Eriocactus leninghausii. Chemotropismus se projevuje růstem kořenů ve směru k zásobě živin.
Otevírání květu při změnách tepla, světla, a vlhkosti či pohyb blizna a prašníků po podráždění jsou další druhy pohybu rostlin (tzv. nastie). Nejnápadnější pohyby má mimosa – citlivka. Ale ani po uhynutí nejsou rostliny zcela nehybné. Dochází k jejich sesychání, pokroucení těla nebo jeho části. Příčinou jsou fyzikální pohyby.
Disimilace (dýcháni) je opakem fotosyntézy. Rozkladem cukrů dochází k uvolňování energie. Ta pak slouží k úhradě energie na životně důležité růstové a rozmnožovací pochody.
Morfózy jsou reakce projevující se změnou intenzity růstu. Např. vlivem ultrafialového záření se růst zpomaluje – trpasličí vzrůst, zmnožuje se otrnění vlna apod. Poranění rostliny může mít za následek bujnější, ale i pomalejší růst postižené části těla. Poškození vegetačních vrcholů vede k deformaci těla.
Dědičnost
Řada kaktusářů se zabývá křížením rostlin za účelem získání neobvyklých nebo lepších vlastností u hybrid. Řeřichovi Mendelovi (1882 – 1884) se prvně podařilo odhalit zákonitosti dědičnosti některých znaků a na jeho počest pak byly pojmenovány jako Mendelovy zákony. K jejich pochopení je třeba nejprve vysvětlit několik základních pojmů.
Dominantní a recesivní znaky: jde o vlastnosti (např. barvu květu), který se může projevit různými znaky (červená – bílá). Jeden z nich – A (např. červená) však vždy převládá (dominuje), kdežto druhý – B se v přítomnosti prvého neuplatňuje (je recesivní). Setkají – li se tyto dva znaky při křížení, projeví se navenek jen dominantní znak (červený květ) a přítomný recesivní znak je potlačen. Hovoříme o fenotypu (podle znaků, které se projevily) a o genotypu (soubor všech znaků – genů, přítomných v genetickém základu rostliny, ať se projevily, nebo jsou v něm přítomny ale jsou potlačeny).
Má – li rostlina pouze jednu dvojici stejných genů (2 x A nebo 2 x B), nazýváme ji homozygotní. Jsou – li geny odlišné (A i B), hovoříme o heterozygotní rostlině. Při schematickém znázorňování pravidel křížení označuje se dominantní znak velkými písmeny a recesivní znak malými písmeny.
V 1. Mendelově zákoně se praví že zkřížíme – li mezi sebou dva homozygotní jedince, lišící se mezi sebou jen jediným znakem, vznikne dceřinná generace – F1 (z latiny filia = dcera), obsahující naprosto stejné hybridy Ve schématech se pro rodičovské rostliny používá označení P1 ( z latiny parent = rodič).
P1: čč x bb Tyto znaky se rozštěpí na čč x bb
1. Schéma křížení
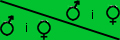 |
č | č |
| b | čb | čb |
| b | čb | čb |
- symbol pro dárce pylu (muž) - symbol pro příjemce pylu (Ž)
č = červená barva květu, recesivní znak
b = bílá barva květu, recesivní znak
čb = růžová barva, protože ani č, ani b nejsou dominantní znak
Jiná situace však bude, je – li jeden znak dominantní: P1 : ČČ x bb
2. Schéma křížení
|
Č | Č |
| b | Čb | Čb |
| b | Čb | Čb |
Č = dominantní červená barva
b = recesivní bílá barva
Čb = červená barva květu, protože červená převládá nad bílou.
V prvém příkladě má dceřinná generace (F1) vždy růžovou barvu, která je kompromisem mezi geny pro červenou a bílou barvu. Nezáleží přitom zda jako dárce (příjemce) pylu použiji rostlinu se znakem č nebo b. Je – li však jeden znak dominantní. (viz příklad č. 2), je sice všechno potomstvo v prvé dceřinné generaci 1 stejné, ale projeví se jen dominantní znak – červená barva květu. Fenotypicky bude mít tedy dceřinná rostlina květ červené barvy. V jejím genetickém základu (genotypu) však bude uchován i znak – gen pro bílou barvu květu. To má význam pro formulaci druhého a třetího Mendelova zákona.
Druhý Mendelův zákon nám umožňuje poznat a získat homozygotní rodičovské rostliny i tehdy, když neznáme opylovače. Praví se v něm, že zkříží – li se mezi sebou hybridy prvé dceřinné linie (F1), vznikne druhá dceřinná linie (F2), v níž se sledované znaky projeví v poměru 1 : 2 : 1 (jsou – li oba dominantní nebo recesivní), nebo v poměru 3 : 1 (je – li jeden dominantní).
Mají – li např. obě rodičovské rostliny z 1. dceřinné generace recesivní znaky (čili F1 = čb x čb), pak platí toto schéma:
3. Schéma křížení
|
č | b |
| č | čč | čb |
| b | čb | bb |
| Znaky: | čč | čb + čb | bb |
| Výsledkem je genotyp: | 1 : 2 : 1 | ||
| I fenotyp je: | 1 : 2 : 1 | ||
Je – li však jeden znak dominantní povahy (Č), platí pro rodičovský pár prvé dceřinné generace (F1 = Čb x Čb) že:
4. Schéma křížení
|
Č | b |
| Č | ČČ | Čb |
| b | Čb | bb |
| Znaky: | ČČ | Čb + Čb | bb |
| Výsledkem je genotyp: | 1 : 2 : 1 | ||
| Ale fenotyp je: | 3 : 1 |
||
neboť dominantní znak Č potlačí recesivní b.
Vždy je tedy možné, že ve 2. dceřinné generaci se objeví homozygotní rodičovský pár (ČČ nebo bb). Jak však najít v příkladu uvedených 3 červenokvětých rostlin tu, která je homozygotní od zbylých dvou hybrid (ČČ od Čb+Čb)? Pomáháme si opět jejich křížením se zjištěnou rodičovskou rostlinou nesoucí recesivní znak bílého květu (bb). Je – li vybraná testovaná rostlina homozygotní, musí podle 1. Mendelova zákona vzniknout čtyři stejné rostliny kvetoucí červeně (viz schéma 2). Nebude – li žádný znak dominantní, pokvetou všechny rostliny růžově (schéma 1.). Není – li však testovaná rostlina homozygotní, (F2 = Čb x bb) nebo (F2 = bb x čb) dojde ke vzniku generace F3:
5. Schéma křížení
|
Č | b |
| b | Čb | bb |
| b | Čb | bb |
6. Schéma křížení
|
č | b |
| b | čb | bb |
| b | čb | bb |
V tomto případě se projeví hybridní původ testovaných rostlin tím, že pokvetou dvěma odlišnými barvami (Čb = červená a bb = bílá, nebo čb = růžová a bb = bílá). Tomuto druhu křížení, kterým lze zjistit zda testovaná rostlina je kříženec, se říká zpětné křížení.
Až dosud jsme se zabývali dědičností pouze jedné dvojice znaků. V životě se však s tak jednoduchými situacemi nesetkáváme. Jak je tomu při dědičnosti dvou párů znaků?
Dědičnost je založena na předávání genů umístěných na chromozonech v jádře buňky. Geny jsou zde vždy uspořádány do skupin označovaných jako alely. Při vzniku pohlavních buněk se alely štěpí, takže zárodečné buňky obsahují geny pouze jednoho partnera, jsou tedy vždy homozygotní.
Křížíme–li mezi sebou jedince, lišící se od sebe dvěma různými alelami, dochází ke vzniku hybrid s různou kombinací alel podle druhého Mendelova zákona.
Praktické využití dosavadních výkladů si můžeme osvětlit na příkladu: chceme zkřížit mezi sebou dva kaktusy, z nich jeden kvete červeně (Č = dominantní znak) a má dva krátké trny (o = recesivní znak), kdežto druhý kaktus kvete žlutě (ž = recesivní znak) a má dlouhé trny (O = dominantní znak). Rodičovské rostliny tedy mají tyto dvě dvojice genů: Ččoo x žžOO. Jejich pohlavní buňky (pyl a zárodek semene) obsahují Čo nebo žO. Podle prvého Mendelova zákona bude prvá dceřinná generace F1 vypadat takto:
7. Schéma křížení
|
Čo | Čo |
| žO | ČožO | ČožO |
| žO | ČožO |
ČožO |
To znamená, že v prvé dceřiné generaci budou pouze červeně kvetoucí, dlouhotrnné hybridy. Alély se však mohou kombinovat zcela nezávisle na sobě, takže vznikají další kombinace genů: ČO, Čo, žO, žo. Křížení bude tedy ve skutečnosti vypadat takto:
8. Schéma křížení
|
ČO | Čo | žO | žo |
| ČO | ČČOO | ČČOo | ČžOO | ČžOo |
| Čo | ČČOo | ČČoo | ČžOo | Čžoo |
| žO | ČžOO | ČžOo | žžOO | žžOo |
| žo | ČžOo | Čžoo | žžOo | žžoo |
Vznikne tedy 16 možných kombinací. Na úhlopříčce vedoucí z levého horního rohu do pravého dolního rodu jsou pouze homozygotní rostliny. Pro pokusníky vznikají tak zajímavé kombinace, lišící se podstatně od fenotypu mateřských rostlin, např. kaktusy s červenými květy a dlouhými trny, nebo žlutě kvetoucí hybridy mající krátké trny.
Výše uvedený příklad je vysvětlením 3. Mendelova zákona, který je označován také jako vznik nových kombinací genů.
Zastoupení čtyř možných fenotypů je zde v poměru 9 : 3 : 3 : 1. Ze schématu vyplývá, že 9 rostlin pokvete červeně a bude mít dlouhé trny, tři hybridy pokvetou červeně a budou mít krátké trny, jiné tři pokvetou žlutě a ponesou dlouhé trny a pouze jedna žlutě kvetoucí hybrida bude mít krátké trny.
Zpravidla ovšem vyséváme větší počet semen, v nichž jsou jedinci s příslušnou kombinací znaků zastoupeni ve výše uvedeném poměru. Menší odchylky od tohoto pravidla samozřejmě nevyvrací jeho obecnou platnost
Řada kaktusářů se zabývá křížením rostlin, velmi časté jsou hybridy astrofyt. Nabízena je celá paleta hybrid vzniklých náhodným křížením, podle zásady „co kvete – spráším“. Cílený výběr rodičů a ještě více žádaných hybrid umožňuje spolehlivěji a rychleji dospět k žádanému cíli – rostlině nových, lepších vlastností. Nezapomínejme, že A.V.Frič byl nejen velkým lovcem kaktusů, ale i velmi zdatným pokusníkem, z jehož rukou vyšla celá armáda hybrid. Věčná škoda, že se mnohé již nedochovaly.
Není hanbou ani chybou přispívat ke vzniku hybridy. Je však naprosto nezodpovědné, vydávat hybridu za „pravý“ nebo „nový“ druh a neuvést jména rodičovských rostlin.
Přehled substrátů
Písek: nejčastější minerální substrát. Sestává ze zrn o velikosti 0,2 – 2 mm, převážně z křemene anebo z jiných zvětralých hornin. Hrubší zrno (od 2 – 20 mm) se označuje štěrk. Čistý písek snadno propouští vodu, neslepuje se, nemá téměř žádné živiny a je lehce kyselý.
Ze zvětralých hornin má pro nás význam:
Pískovec: usazeniny vzniklé vazbou pískových zrn pomoci kyseliny křemičité a železa. Má zpravidla kyselou reakci, je chudý na živiny, obsahuje však stopové prvky.
Granit: je prahorninou, nachází se obvykle jako shluky (konglomeráty) živce, slídy, křemene je slabě kyselý.
Bazalt: reaguje zásaditě, je sopečnou vyvřelinou sestávající z různých látek. Je tvrdý, těžký nepřijímá vodu a velmi pomalu zvětrává. Moučky z bazaltu se užívá ke zlepšení kyselých půd, obsahuje živiny a stopové prvky.
Porfyr: rovněž stará vyvřelina, velmi hustá s jemnými zrny a živinami, obsahuje usazeniny draselných solí.
Diabas: patří do skupiny starých vyvřelin. Je zelenočerný, má zrnitou strukturu. Zvětralý písek z něj je lehce kyselý, výživný, lépe uchovává vláhu než předchozí vyvřeliny.
Pemza: je sopečný písek často užívaný ke stavebním pracím pro lehkost a tepelnou izolaci. Je porézní, lehký, obsahuje různé minerály. Často má zásaditou reakci, proto se doporučuje jej před použitím dobře vyprat. Téměř neobsahuje živiny, je silně vodopropustný, dobře uchovává vlhkost a provzdušňuje půdu.
Lavalit: používá se teprve nedlouho, pochází z horkého sopečného popele prostoupeného plynovými bublinám. Je porézní, lehký, červenohnědé barvy. Neobsahuje žádné volné živiny, zadržuje vodu a provzdušňuje každý substrát.
Slín: je usazenina skládající se převážně z vápence a jílu. Na vzduchu se rychle rozpadá, je chudý na živiny, má zásaditou reakci. Slínovité půdy špatně propouští vodu a jsou chladné, pro kaktusy nejsou příliš vhodné.
Spraš: je velmi kyprá, jemnozrnná zemina, velmi úrodná o pH 7 až 7,5, sestávající ze slínu, jemných zrn křemičitanů, vápence a zbytků prahorních živočichů.
Jemnozrnné substráty jsou hlína a jíl. Jíl je velmi těžký a využívá se hlavně k výrobě kameniny.
Hlína: má velmi jemnou strukturu danou pravěkými usazeninami. Pokud zcela nevyschne je vhodná jako přídavek do substrátů pro kaktusy. Její pH může být kyselé i zásadité. Snadno předává kořenům živiny a dlouho udržuje vláhu.
Vápenec: má mezi minerály zvláštní postavení. Rozpouští se ve vodě a vyvolává v ní i v půdě zásaditou reakci. Uvolňuje v půdě přítomné fosfáty které se tak stanou využitelné pro rostliny. Vápník je jednou z hlavních živin rostlin. Většina kaktusů vápník potřebuje. Protože však současně obvykle vyžadují kyselý substrát, podáváme jim vápník v podobě síranu vápenatého, tj. sádry. Sádra má kyselou reakci a v přírodě se nachází ve slínu, alabastru a jílu. V podobě křídy, mramoru, argonitu, lomitu a jiných hornin je vápník obsažen ve vazbě na uhlík, nejčastěji v podobě uhličitanu vápenatého. Sádru nelze přímo přidávat do půdy. Ztvrdlé kousky sádry roztlučeme na kousky a přidáme po 1 – 2 na dno květináčů.
Křemeny: jsou oxidy křemíku obsažené v písku a také v polodrahokamech. Vhodné k obkládání krčků choulostivých rostlin.
Jíl: vzniká zvětráváním jílovitých silikátů – křemičitanů. Ve vodou chudých oblastech bývá často prostoupen různými solemi.
Antuka: dříve často doporučovaná k pěstování kaktusů, má již blíže k tzv. chemickým substrátům. Je sterilní, obsahuje jen minerální součásti, podporuje růst kořenů, zadržuje vodu a je vzdušná. Reakci pH lze upravit přídavkem hlíny nebo jílu.
Kerazmit: jsou zhruba kulaté jako cihla vypálené kousky jílu užívané ve stavebnictví a zahradnictví. Jsou vhodné pro hydrokulturu, dobře udržují vzduch a vodu. Působí také dekorativně.
Čistě umělými – chemickými substráty jsou:
Perlit: je to bílý drobivý perlivý granulát křemičitanu hliníkového, velmi lehký, teplý, neutrální reakce, vhodný i pro choulostivé rostliny. Dobře si uchovává svoji strukturu, zadržuje vodu a vzduch. Je dobrou příměsí k substrátům pro kaktusy.
Hygromull: Jsou to měkké vločky dobře si uchovávající tvar, lehké. Pomalu se rozkládají a poskytují výživu kořenům. Užívá se jako příměs k substrátům až do 12 objemových %.
Vermikulit: je lehký, korku podobný granulát užívaný k tepelné izolaci ve stavebnictví. Vhodný jako příměs provzdušnění jílů a těžkých půd.
Humusními půdami rozumíme všechny substráty obsahující rozpadlé organické látky. Patří mezi ně:
Kompost: je to velmi koncentrovaný humus získaný řízeným rozkladem organických zbytků. Biologicky je velmi bohatý a, podle druhu a složení organických látek, velmi výživný, s obsahem velkého množství dusíku. Pro vysoký obsah mikrobů není pro většinu kaktusů vhodný. Stará kompostní zem má již mikrobů méně a lze ji použít pro rostliny pocházejících z vlhkých oblastí.
Listovka: je humusovitou, mikrobně chudší půdou, vhodnou pro všechny kaktusy nevyžadující čistě minerální substrát. Obsahuje málo dusíku a má drobtovitou strukturu.
Zvětralé jehličí: obsahuje méně humusu, ale dost éterických pryskyřic.
Bukovka: je kyselá a obsahuje pro rostliny nevhodnou tříslovinu.
Humus a kompost vznikají jen za přístupu vzduchu a dostatečné vlhkosti (ne však přemokření). Pochází ze zelených částí rostlin rozkládajících se způsobem saprofytních bakterií. Takto dochází v přírodě k opakovanému využití odumřelých organických látek. Podílí se na něm vše živé – člověk, zvířata, rostliny, mikroby – a sami se stávají oběťmi tohoto koloběhu. Pokud se do něj zasahuje zbytečným pálením organických látek, přerušujeme tento oběh živin se všemi biologicko – ekologickými důsledky.
Humus nevzniká tam, kde chybí vlhko (pouště). Slunce a vítr brání vzniku humusu, naopak zvířecí výkaly rozklad urychlují (pokud v nich nejsou přítomny tuky). Při nízkých teplotách se většina bakterií nemnoží, takže rozklad organických látek se na čas zastavuje. Během rozkladu organické hmoty se uvolňuje teplo, takže kompost je teplejší než jeho okolí (tzv. pařeništní efekt).
Na pobřežních horách Peru a Mexika či Dolní Kalifornie, kde žijí obrovská množství ptactva, roste mnoho kaktusů v silných vrstvách humusu. Chybí zde však voda, která by humus učinila pro rostliny využitelný.
Čím sušší je krajina vystavená slunci a větru, tím méně v ní může existovat rostlin. Odumřelé rostliny rychle vysychají a nepodílí se pak na tolik potřebném vzniku humusu.
Pouštní půdy ztrácí více vody odparem, než kolik jí přijmou v podobě deště nebo rosy. Jejich povrch je proto často značně prosycen minerálními solemi. Rostliny pocházející z těchto končin proto lehce snáší přihnojování minerálními solemi, ale jsou citlivé na větší příměs humusu v půdě. Zvláště pak dojde–li po větší zálivce k bohatšímu mikrobiálnímu životu v humusní půdě, odumírají jim choulostivé kořeny. Rostliny pocházející z vlhčích oblastí si snadněji zvykají na půdy s nižším obsahem humusu než pouštní rostliny na humus. Tento poznatek je základem kaktusářské praxe!
Rašelina: má zvláštní postavení mezi organickými látkami humusovitého charakteru. Během tisíciletí nashromážděné za nepřístupu vzduchu rostlinné hmoty, které se nerozpadly, ale spíše konzervovaly, mají svou povahou poměrně blízko k povrchovému hnědému uhlí. Často nabízená světlá rašelina ze svrchních, mladších vrstev přijímá snadněji rostliny a mikroby než starší černá rašelina. Pro vysoký obsah huminových kyselin a kyseliny uhličité má rašelina silně kyselou reakci (pH 2 – 5). Usnadňuje rozpouštění živin z půdy a umožňuje jejich využití rostlinám. Pomocí samotné rašeliny lze zásadité půdy okyselit. Rašelina je především stimulátorem života v půdě, nikoliv hnojivem! Zásoby rašeliny jsou na světě téměř vyčerpány. Občas se ještě nabízí rašelinové směsi a rašelina obohacená o živiny, která je zvláště vhodná pro epifyty.
Komerčně nabízené půdní směsi: Využívají hlavně vydatného přídavku umělých substrátů, jejich předností je nízká váha, velká pórovitost a nasáklivost. Zvláště při zálivce podmokem je tato nasáklivost nezbytná. V současné době nabízené univerzální substráty obsahují hlavně jíl (asi 40%) a přídavek rašeliny k provzdušnění a zvýšení objemu. K této základní směsi se přidává buď uhličitan vápenatý (k okyselení), nebo jen hnojivo. Obvykle se přidává 12% dusíku, 13% fosforečnanů, 17% draslíku, 1% hořčíku a stopové prvky, hlavně pak železo, mangan, měď, bor, zinek a molybden.
Na závěr ještě radu: Starou zem po přesazování nevyhazujte. Před opakovaným použitím však musí být sterilizována. Lze k tomu užít starý 10 litrový hrnec s přiléhavou pokličkou. Hrnec asi ze 3/4 naplníte použitou zeminou (vhodnější je však do hrnce vložit mikrotenový sáček se zeminou!), přilijete 2 – 3 litry horké vody a necháte 25 – 30 minut na ohni propařovat. Nato zeminu přesypte do kovové misky nebo ji v sáčku vytáhněte a sáček pak nechejte nahoře široce otevřený. Za 3 – 7 dnů je možné sterilizovanou zem opět použít, případně k ní přidat trochu čerstvé půdy k rychlejšímu oživení půdní mikroflóry. Staré organické látky se po sterilizaci rozpadly, zem je zcela minerální a neobsahuje žádné škůdce.
Nové poznatky o kaktusových rostlinách
Dle práce N. H. Bokeho „Developmental Morphology and Anatomy in Cactaceae“, BoiScience, vol. 30, č. 9 / 1979, přeložil a zkrátil Dr. Plesník.
Kaktusy hned od svého objevení vzbudily zájem jak amatérů, tak profesionálních botaniků. Podrobnější znalosti o stavbě jejich těla (anatomie), o jejich vývoji a srovnávací studie s jinými rostlinami jsou jen útržkovité, neboť v tradičních centrech botanického výzkumu lze jen stěží získat dostatečný počet potřebných vzorků materiálu. I dnes lze spočítat na prstech jedné ruky vědce, kteří se zabývají asi 150 rody a 2000 druhy rostlin, patřící do čeledi Cactaceae. Přesto stále přibývá nových poznatků.
Pro zvláštnosti stavby těla a květů jsou potíže se zařazením kaktusů mezi ostatní kvetoucí rostliny. Podle poznatků embryologie (nauka o vývoji zárodku), struktury květů a těla patří čeleď kaktusovitých do řádu Caryophylales. Tomu nasvědčuje přítomnost betakyaninů, téměř ve všech (mimo 2) čeledích, zařazených do tohoto řádu. Betakyaniny jsou purpurové až červené pigmenty obsahující ve své molekule dusík. Vyznačují se dobrou rozpustností ve vodě. Mnohem rozšířenější anthokyaniny dusík v molekule nemají. Typickou barvu červené řepy tvoří betakyanin (převzato z angličtiny, kde beet = řepa). U kvetoucích rostlin je toto barvivo vzácné. Chybějící důkazy o prapředcích kaktusů mohou být samy o sobě znamenat, že původ kaktusů není tak dávný. Čeleď Aizoaceae, která se obvykle považuje za blízce příbuznou kaktusům, se vyvíjela zcela odlišným způsobem jako listnaté sukulenty. Podobnost jejich květů květům kaktusů je jen povrchní. Obdobný vývoj a struktura semeníku a plodů u řady afrických sukulentů s plody Pereskia pititache patřící mezi tropické, olistěné kaktusy, je příkladem souběžného vývoje (paralelismu). To svědčí jak pro společného prapředka, tak pro vývoj za podobných vlivů prostředí.
Výhony, cévy a žlázy kaktusů
Olistěné kaktusy tropických lesů jsou nepochybně nejméně přizpůsobeny životu kaktusů. Tím, že mají široké listy, vznikající z typické listové zárodečné tkáně (meristému), že mají zřetelná internoida (části osy rostliny mezi místy, na něž přisedají listy) a jen slabou sukulenci, mohou povrchnímu pozorovateli připomínat kaktusy jen svými trny. I když pereskie nejsou sukulenty, mají řadu jejich znaků, značný přírůstek obvodu primárního cévního svazku a průměr dřeně. V menší míře se s tímto jevem setkáváme u všech pereskií, velmi výrazné jsou tyto znaky u ostatních příslušníků čeledi kaktusovitých. Velké pereskie mají kmen o průměru 30 – 40 cm s dření dosahující 5 – 6 cm tloušťky. Chybí jim vnitřní dřevitá tkáň, kterou nemá žádný kaktus. Zvláštnosti těla kaktusů jsou mimo obvyklého cévního svazku ještě dobře vyvinuté a vzájemně propojené systémy cév, uložených pod pokožkou (kortikální) a v dřeni (medulární). Nalézají se u řady cereusů, jinde však ne. I když hodně druhů má systém podkožních a dřeňových cév (koryfanty, echinocereusy, dolichotele) a některé mají jen podkožní cévy (část mamilárií), žádný kaktus nemá pouze dřeňové cévy.
I když se ví, že kaktusy mají buňky a tkáně tvořící hlen, byly tyto struktury studovány dosud jen zřídka. Hlenotvorné buňky jsou zvláště časté u opuncií a echinocereusů. Hlenovité kanálky a dutinky jsou velmi hojné u Peireskia pititache a u ariokarpusů mění se jejich obsah na hlenovitou hmotu, v níž jsou přítomny volné buňky i jejich shluky.
Mlékovody, obsahující mléčnou nebo polomléčnou šťávu (latex), mají některé mamilárie a příbuzné rody. Mauseth při podrobném studiu těchto rostlin zjistil, že mlékovody bývají přítomny v dřeni, pod pokožkou, tedy v tzv. kortexu a v hrbolech. Chybí však vždy v květech a také u semenáčků mladších osmi měsíců. Vznikají až ve starších tkáních, nikdy se netvoří v zárodečné tkáni (meristému) a v jeho okolí.
Meristém - zárodečná tkáň
Mezi krytosemenými rostlinami mají právě kaktusy v temeni uložený (apikálně uložený) největší meristém. Dalo by se to očekávat u obrovitých rodů, např. Echinocactus grandis, u nichž epikální kužel zárodečné tkáně může mít v průměru až 2 mm. Ale i některé mnohem drobnější echinocereusy, mamilárie, koryfany mají apikální meristém o průměru 1 – 1,5 mm. U kaktusů se silným a výrazně žebrovaným tělem (Carnegia, Echinocactus), řidčeji také u menších (Echinocereus) je apikální meristém uložen v dutině zaplněné vlasy a trny, vzniklé růstem žeber do stran. Pokud není zničena nebo poškozena funguje u většiny kaktusů tato zárodečná tkáň až do konce jejich života. U opuncií je aktivní jen určitou dobu, zpravidla jen jedno vegetační období, Další růst rostliny je pak dán pučením z axilárních pupenů. Proto se tělo opuncií skládá z jednotlivých článků. Jsou – li články ulomeny větrem nebo zvěří, vytváří vlastní kořeny a vyrůstá samostatná rostlina.
Kaktusy které přirozeně nikdy neodnožují, nebo jen velice zřídka, lze k tomu donutit odstraněním temene. Odnože se pak tvoří z aktivizovaných axilárních pupenů. Nejméně dva rody – Obregonia aStrombocactus – za normálních podmínek nikdy neodnožují a nevytvoří postraní výhony ani když jim odstraníme temeno. Přídatné pupeny jsou však uloženy na řezné ráně, výhradně v okolí cévního svazku.
Pravá dichotomie (rozpůlení růstového vrcholu na dvě poloviny) je mezi dvouděložními rostlinami vzácností. U kaktusů se nejčastěji objevuje mezi mamiláriemi. Dochází tak ke vzniku skupiny hlav zhruba stejných rozměrů. Jiné trsovitě rostoucí mamilárie tvoří nové hlavičky běžným odnožováním.
Listy hrboly a areoly
U mnoha dvouděložních rostlin vyrůstají listy z více méně vyčnívajícího hrbolu. Ten bývá označován jako listová báze a vyrůstají z ní, jsou – li přítomny, také palisty. Němci je označují jako “Unterblatt“ na rozdíl od „Oberblatt“, sestávajícího z listové čepele a listové osy – petioly. Při svém vývoji kaktusy využily báze a vyjma peireskií a opuncií zredukovaly čepel i na osu listu na nepatrné zbytky. Listová báze se změnila na různé typy hrbolů. Povrch kaktusů tvoří navzájem se stýkající hrboly nebo žebra. Hranice mezi listem a bází není zvlášť zřetelná. U opuncií a peireskií je vyznačen rýhou na spodní straně.
I když všechny kaktusy mají zárodky (primordia) listů, pouze peireskie a opuncie tvoří listy podobné listům jiných rostlin. Mnoho dalších kaktusů má drobounké listy jen v okolí apikálního meristému. Listy na probouzejících se pupenech mohou být mnohem větší než na vyzrálých stoncích, jak je tomu např. u rodu Bartschella. Ale i u opuncií dochází již v prvé růstové sezóně ke ztrátě listů. Vyjímkou je jenOp. subulata mající největší lístky ze všech opuncií.
U některých kaktusů nabývají hrboly teprve druhotně listového tvaru (Ariocarpus, Leuchtenbergia, Obregonia). Jiné tvoří žebernatý nebo plochý výhon vznikající tím, že listové báze ležící v jedné vertikální linii rostou více nebo méně intenzivně. Zygocactus a Epiphyllum mají výhony podobné listům proto, že jejich žebra byla redukována jen na dvě.
Zvláštním růstovým útvarem kaktusů je areola, což je axilárně uložený pupen (axila = úhel mezi osou stonku rostliny a osou odnože, listu apod. vyrůstající z těla rostliny, u kaktusů je to prohlubenina mezi hrboly nebo části hrbolů), z nějž vyrůstají trny a nebo tenké a malé trny označované jako glochidy. Navíc odtud vyrůstají i vlasy. Z areol, podobně jako z axilárních pupenů, mohou vznikat odnože nebo květy. Mohou zůstat v klidu nebo mohou stále tvořit trny či glochidy, což vidíme u některých velkých opuncií.
Většina kaktusů má různé typy trnů: dlouhé – krátké, podobné vlasům, papírovité, háčkovité, ploché, silné, tenké, zpeřené hladké nebo žláznaté. I tyto rody kaktusů které v dospělosti trnů nemají (Ariocarpus, Lophophora, Rhipsalis) mají trny jako klíčenci. Zárodky trnů pochází z meristému stejně jako zárodky listů, nemají však cévy. Růst trnů závisí na délce aktivity a množství dělících se buněk, které rozhoduje o délce, síle a vzájemném postavení trnů. Rozdíly v délce buněk na špičce trnu vedou ke vzniku háčků typických pro některé druhy. Trny se zpětnými háčky tj. jednotlivými povrchovými buňkami rostoucími směrem k tělu, mají jen opuncie, peireskiopsis a příbuzné rody. Vyzrávání trnů je provázeno jejich značným zpevněním (sklerifikací), při čemž všechny trny na areole jsou svými basemi více či méně spojeny a pevně drží na rostlině. Styk s glochidami a trny Op. bigelovii či Op. tunicata je zvlášť nepříjemný, neboť glochidy se snadno zachycují všeho, co přijde do jejich blízkosti. U jiných kaktusů se část trnů mění na aktivně vylučují žláznaté útvary (Ferocactus, Hamatocactus, Coryphanta, Toumeya). Rozložení trnů na mladých areolách je obvykle vzhledem k meristému excentrické. Většina trnů je na straně vzdálenější od osy těla. Glochidy zpravidla vyrůstají na straně bližší od těla.
Většina kaktusů má všechny areoly stejné, lokalizované na hrbolu. Každá areola má vlastní zárodečnou tkáň, z níž vznikají trny, odnože nebo květy. Jejich tvar je okrouhlý až oválný jsou uloženy v malé proláklině na vrcholku hrbolu. U některých koryfant jsou areoly silně protáhlé. Rýhy na vrcholech dospělých rostlin dosahují skoro až do axil. Proto se zdá jako by květy a odnože vyrůstaly z axil, kdežto trny z vrcholku hrbolů. Zárodečná tkáň trnů je někdy uprostřed délky rýhy.
Hrboly (tuberkula), nesoucí dvě odlišné areoly, má jen podřád Cyryphanthanae (mamillárie a příbuzné rody) a několik obvykle řazených mezi Echinocactanae (Ariocarpus, Epithelantha, Pelecyphora). U těchto rostlin došlo k rozložení zárodečné tkáně areoly na k tělu přilehlé straně vznikajícího hrbolu. Následkem vyzrání tkáně uprostřed hrbolu došlo k odizolování zárodečné tkáně pro trny (na špičce hrbolu) od tkáně axilách (odnože, květy). Rýha je téměř neznatelná, nebo zcela chybí. Tato dvojakost (dimorfismus) areol (rozdělení areoly na dvě části) není stále ani v rámci příbuzných rodů, ani uvnitř druhů. Ač areoly Cor. vivipara jsou výhradně monoformní, areoly Cor. erecta mohou být obojího typu: na rostoucích hrbolech jsou monoformní a mohou z nich vyrůst květy či odnože, na vrcholech bez rýhy jsou areoly dimorfní, při čemž axilární merestém vždy dává vznik jen květům. Pouze u epitelant jsou obě části areoly umístěny na vrcholu hrbolu.
Kořeny a květy
Ve srovnání s výhony jsou kořeny kaktusů zkoumány poměrně zřídka. Kořeny snadno vznikají z areol na krčcích článků opuncií, které přijdou do styku s půdou. Vzdušné kořeny vyráží z výhonů poléhavých nebo převisle rostoucích druhů.
Květy kaktusů jsou jedinečné díky souboru charakteristických znaků: Početným okvětní plátkům, které se jen nevýrazně člení na zevní (sepály) a vnitřní (petály), řadou spirálovitě postavených tyčinek, pestíkem skládajícím se ze 4 a více navzájem spojených částí obklopených kalichem květu atd. Přítomnost listů, trnů nebo šupin na semenících mnoha kaktusů a dlouhé květní trubky např. u echinopsisů či selenicereusů naznačují, že semeníky jsou pokryty tkání stonku. Největší rozdíly mezi květy kaktusů a květy rostlin byly nalezeny ve vzniku a složení pestíku a jeho částí. Podobnou stavbou semeníku jako kaktusy má jen rod Mesembryathemum. Podle Buxbauma a dalších, jsou kaktusy a mesembryatémata dvě samostatně se vyvíjející čeledě s podobným průběhem vývoje.
Ekologie
Adaptace prvotních kaktusů na sucho nejspíše probíhala na skalnatých výběžcích vlhkých tropických pralesů odkud kaktusy pochází. Postupně nabyly schopnost obsazovat stále sušší oblasti jihu a severu. I když většina kaktusů se přizpůsobila suchomilnému životu, malá část si ponechala ploché, listům podobné výhony, zpravidla nemá trny a vrátila se z pouští zpět do pralesa jako epifytní druhy (Epiphyllum, Zygocactus). Epifytní růst přináší podobné problémy se získáváním vody jako růst na skaliskách. Pro zpětnou migraci do pralesa svědčí tvar jejich semenáčků: čtyř – až mnohostranný ostnatý válec. Tento tvar si řada cereusů uchovala až do dospělosti. Chybění listů a silné ztlustění zevních tkání stonku podstatně omezilo ztrátu vody výparem. Z toho plynoucí ometení plochy pro fotosyntézu má však za následek pomalý růst kaktusů. Proto kaktusy nemohou čelit konkurenci jiných rostlin v prostředí, umožňující růst i jiných rostlin. Jejich naleziště bývají ohraničena na nehostinné lokality. Protože se kaktusy přizpůsobily suchu tím, že střádají vodu do zásoby, nikoliv však odolností proti vyschnutí, bývají jejich nejbohatší naleziště v teplých, poměrně vlhkých pouštích. Je třeba, aby deště se opakovaly dostatečně často k úhradě ztráty vody v době, kdy teplota půdy umožňuje vstřebání vody. Deště v chladném období jsou nepříznivé.